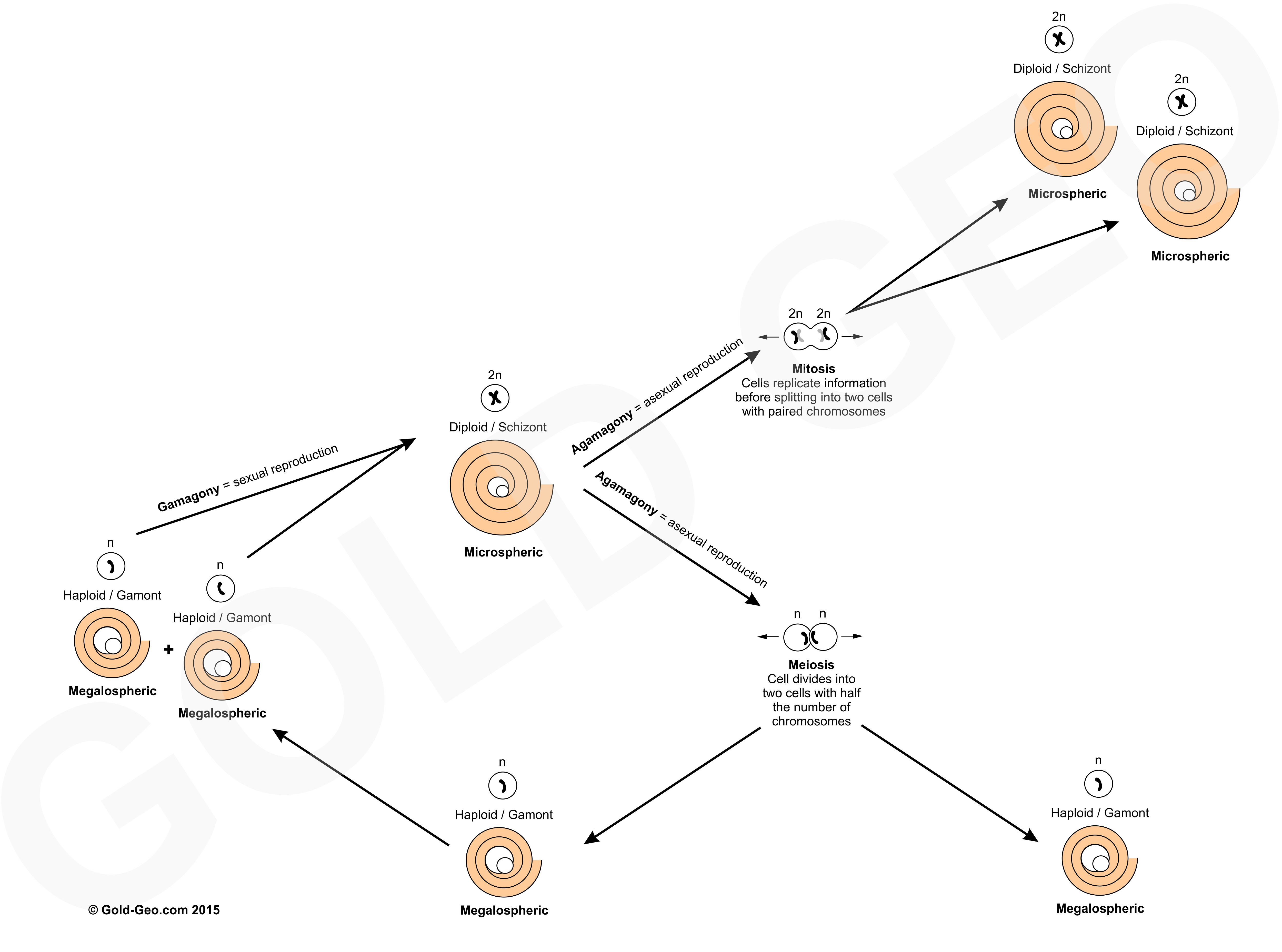
Why do coral reefs, which are home to a high diversity of life, thrive in such nutrient-deficient (oligotrophic) waters? This ‘nutrient paradox’ is a question that has puzzled scientists since Charles Darwin (1842) observed this phenomenon during his voyage on the HMS Beagle. This is also true of foraminifera where a large number of species are also found in oligotrophic settings. So how do nutrients effect the life cycle and distribution of foraminifera? To answer this, we must first define what we mean by nutrients and how these determine marine trophic zones.

Nutrients primarily refers to fixed nitrogen (NH4+, NO2–, NO3–), phosphate (PO43-), iron (Fe2+, Fe3+) and silicate (SiO32-, SiO44−) ions. These are essential nutrients needed in small amounts by photosynthetic organisms to metabolise their cells and are used for cell growth, maintenance and reproduction (Hallock and Schlager, 1986; Hallock, 2001).
Trophic zones may be defined by the amount of chlorophyll from photosynthetic organisms and nutrients within a given environment. In the following examples the amount of phosphate defines the zone:
Low nutrients – Oligotrophic – 0.0-2.6 μg/l chlorophyll, 0.0-12.0 μg/l phosphate
Moderate nutrients – Mesotrophic – 2.6-20.0 μg/l chlorphyll, 12.0-24.0 μg/l phosphate
High nutrients – Eutrophic – 20.0-56.0 μg/l chlorophyll, 24.0-96.0 μg/l phosphate
Phosphate and iron in large amounts is known to inhibit carbonate crystal growth. Phosphate inhibits the growth of both calcite and aragonite, whereas iron inhibits the growth of calcite but not aragonite (van Langerak et al., 1999). The inhibition of crystal growth by trace amounts of ions is caused by the adsorption of the ions onto crystal growth sites (van Langerak et al., 1999). This partially explains why some organisms that precipitate a carbonate (calcite or aragonite) skeleton do not flourish in meso- or eutrophic environments where these nutrients are present.
Another reason why nutrient-rich environments do not favour the growth of benthonic photosynthetic organisms is that with increased nutrients there is an increased abundance of phytoplankton (a phytoplankton ‘bloom’) at the near surface. These phytoplankton occur in large numbers as they take up the nutrients causing them to reproduce and multiply rapidly. This causes the surface waters to become murky and the amount of light reaching the seafloor to reduce – bad news for bottom-dwelling photosynthetic organisms. This scenario also occurs where there is significant terrestrial run-off, such as a river delta, that dumps a large amount of fine-grained material into the shallow marine environment causing increased murkiness and reduced light intensity. However, recent discoveries of coral reefs close to the mouths of large rivers near the Amazon and in Indonesia are challenging this idea.

A further explanation for the nutrient paradox is role of gross primary productivity (GPP) vs net primary productivity (NPP). Primary productivity is the rate at which energy is fixed into organic carbon, such as simple sugars, during photosynthesis (Hallock, 2001). GPP is a measure of the total amount of energy fixed to organic carbon regardless of whether this is used for respiration or cell growth (Hallock, 2001). NPP is the amount of energy fixed during photosynthesis that is available for cell growth which excludes the amount of energy used in respiration (Hallock, 2001). In reefs, GPP can be very high as nutrient-depleted clear waters with increased light penetration result in increased rates of photosynthesis and respiration, whereas NPP is actually very low (Hallock and Schlager, 1986; Hallock, 2001). Therefore, observations of thriving coral reefs in nutrient deficient waters relate to interpretations of GPP rather than NPP (Hallock and Schlager, 1986; Hallock, 2001).

Most calcareous benthic foraminifera are oligotrophic (favour low nutrients), these include nummulitids, lepidocyclinids and orthophragminids. This is because they house photosynthetic algal symbionts within their tests which need clear water to photosynthesise. These foraminifera often reach large sizes, with larger size corresponding to lower amounts of nutrients. These foraminifera grow large as a combination of having to provide a home for a thriving community of photosynthetic symbionts and by having a long life cycle to allow them enough time to gather enough of the required amount of scarcely available nutrients to enable them to reproduce. Some rotaliids, however, can tolerate high nutrients and can be found in mesotrophic settings. In mixed carbonate-siliciclastic (MCS) settings of the Mahakam Delta in East Kalimantan, Indonesia, Novak and Renema (2018) found distinct assemblages containing the genera Lepidosemicyclina, Anomalinella and Pseudorotalia which are restricted to MCS environments with fine-grained, muddy substrates. Small miogypsinids are dominant in shallow water settings with high terrigenous input (Novak and Renema, 2018).
Most miliolids are also oligotrophic and have thick microgranular walls which protect them from harmful UV radiation in shallow, clear water. However, some miliolids occur in deep water where nutrients are also scarce. Other agglutinated foraminifera are opportunistic and can tolerate a variety of stressed environments including increased nutrients and salinity. Therefore a fossil assemblage dominated by agglutinated foraminifera may suggest deposition within a mesotrophic setting. Other organisms that can tolerate high nutrients include bryozoa, echinoids and sponges where silicate is a nutrient for the latter.
References:
Darwin, C.R., 1842. The Structure and Distribution of Coral Reefs: Being the First Part of the Geology of the Voyage of the Beagle, Under the Command of Capt. Fitzroy,… During the Years 1832 to 1836. By Charles Darwin,… Published with the Approval of the Lords Commissioners of Her Majesty’s Treasury. Smith, Elder and Company, 65, Cornhill.
Hallock, P., 2001. Coral reefs, carbonate sediments, nutrients and global change. In: Stanley Jr., G.D. The history and sedimentology of ancient reef systems. p. 388-428
Hallock, P. and Schlager, W., 1986. Nutrient excess and the demise of coral reefs and carbonate platforms. Palaios, pp.389-398.
Novak, V. and Renema, W., 2018. Ecological tolerances of Miocene larger benthic foraminifera from Indonesia. Journal of Asian Earth Sciences, 151, pp.301-323.
van Langerak, E., Beekmans, M.M.H., Beun, J.J., Hamelers, H.V.M. and Lettinga, G., 1999. Influence of phosphate and iron on the extent of calcium carbonate precipitation during anaerobic digestion. Journal of Chemical Technology and Biotechnology, 74(11), pp.1030-1036.